|
|
|
страница 1
Глава 9
АДАПТАЦИЯ МЫШЕЧНОЙ, КОСТНОЙ И СОЕДИНИТЕЛЬНОЙ ТКАНЕЙ
Структурно-функциональные особенности
двигательных единиц мышц
Мышца является исключительно разнородной тканью, состоящей преимущественно из мышечных волокон, соединительнотканных, нервных и сосудистых элементов, которые в комплексе обеспечивают ее главную функцию — активное сокращение. В структуре мышечной ткани различают два типа мышечных волокон — медленносокраща-ющиеся (МС) и быстросокращающиеся (БС).
МС-волокна обладают следующими свойствами: небольшой скоростью сокращения, большим количеством митохондрий («энергоцентр» клетки), высокой активностью оксидативных энзимов (протеины способствуют быстрой активизации источников энергии), широкой васкуляризацией (большое количество капилляров), высоким потенциалом накопления гликогена. БС-волокна имеют менее развитую сеть капилляров, меньшее число митохондрий, высокую гликолитическую способность, высокую активность неоксидативных энзимов и более высокую скорость сокращения (Gollnick, Hodgson, 1986; Noth, 1992).
В одной и той же мышце содержатся БС- и МС-волокна. БС-волокна содержат активный фермент АТФазу, который расщепляет АТФ с образованием больших количеств энергии, что обеспечивает быстрое сокращение волокон. В МС-волок-нах активность АТФазы низкая, в связи с чем энергообразование в них совершается медленно. Ферментативное расщепление АТФ считается одним из важных факторов, определяющих присущую мышце скорость сокращения. Ферменты, которые расщепляют сахар и жиры, активны в МС-волокнах, что позволяет объяснить весьма существенные различия между типами волокон (Эвартс, 1984; Уилмор, Костилл, 2001).
Различают две подгруппы БС-волокон: БСа и БСб. БСа-волокна называют быстросокращающи-мися оксидативно-гликолитическими волокнами. Они отличаются высокими сократительными способностями и одновременно обладают высокой сопротивляемостью утомлению. Эти волокна хорошо подвержены тренировке на выносливость. БСб-волокна — классический тип быстросокраща-ющихся волокон, работа которых связана с использованием анаэробных источников энергии. Каждый из указанных типов волокон достаточно хорошо идентифицируется под микроскопом после соответствующего окрашивания срезов.
Соотношение мышечных волокон разных типов детерминировано генетически. Вероятно, структура мышечного волокна, соотношение волокон различного типа заложены на уровне ДНК и в значительной мере определяются особенностями нейромышечной регуляции, о чем вполне убедительно свидетельствуют исследования, в которых изучалось влияние на изменение типа мышечного волокна перекрестной иннервации (Staron, Pette, 1990). Таким образом, генетически заданный тип иннервации обеспечивает формирование фенотипа мышечной ткани, которая лишь в относительно узких границах может быть модифицирована напряженной тренировкой (Diskuth, 2004).
Мышечные волокна объединяются в двигательные единицы (группы мышечных волокон, иннер-вируемых одним мотонейроном), каждая из которых состоит из мышечных волокон определенного типа. Строение и функции мотонейронов соответствуют строению и функциям объединяемых ими мышечных волокон. Мотонейрон медленносокра-щающейся двигательной единицы объединяет группы из 10—180 МС-волокон и имеет небольшое клеточное тело. Мотонейрон быстросокраща-ющейся двигательной единицы иннервирует от 300 до 800 БС-волокон и отличается большим клеточ ным телом и большим количеством нервных отростков (Энока, 2000).
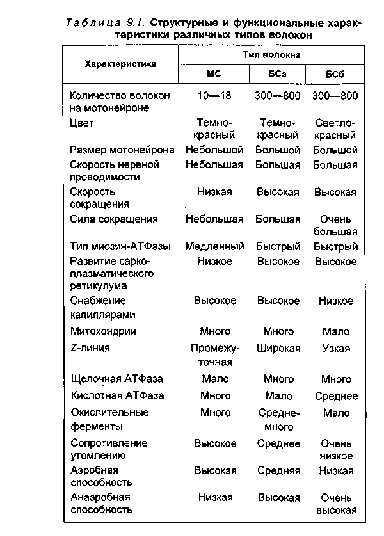
В последние десятилетия в связи с развитием биохимии и морфологии появилась возможность значительно глубже изучить структуру и функции мышечных волокон и двигательных единиц мышц, расширить представления об особенностях их адаптации к тренировочным и соревновательным нагрузкам. Наиболее общие представления о структуре и функциональных возможностях различных типов мышечных волокон приведены в табл. 9.1.
Время, необходимое для максимального напряжения БС-волокон, обычно не превышает 0,3— 0,5 с, в то время как МС-волокна способны развить максимальное напряжение лишь через 0,8— 1,0 с (Мохан и др., 2001). Активность анаэробных ферментов БС-волокон может в два раза и более превышать активность этих ферментов в МС-во-локнах. В то же время активность аэробных ферментов в МС-волокнах примерно в два раза превышает аналогичные показатели БС-волокон (Essen et al., 1975; Gollnick,
Matova, 1984).
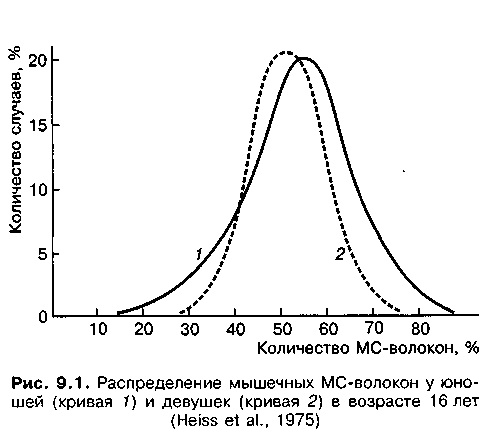
Говоря о пропорциях различных мышечных волокон у человека, следует отметить, что и у мужчин, и у женщин МС-волокон несколько больше (по данным разных авторов — от 52 до 55 %). Это достаточно ярко иллюстрируется результатами исследований, в которых участвовали 45 девушек и 70 юношей в возрасте 16 лет (рис. 9.1). Среди БС-волокон преобладают волокна типа БСа (30—35 %). БСб-волокон значительно меньше — 12—15 %. При этом следует указать, что в мышцах у женщин, в отличие от мужчин, реже превалирует какой-либо один тип волокон (Gollnick et al., 1972; Thorstensson, 1977). Однако результаты отдельных исследований позволяют говорить о том, что определенная часть быстросокращающихся мышечных волокон заложена в человеке, однако подавлена в процессе генотипической и фенотипической адаптации. Эта часть мышечных волокон доступна для генных манипуляций, что вызывает особое беспокойство в свете перспектив в борьбе с генным допингом (Diskuth, 2004).
При рассмотрении адаптации мышц человека в процессе тренировки следует учитывать особенности распределения мышечных волокон различного типа в одной мышце и в разных мышцах. Специальные исследования показали, что в одной мышце изменения в содержании различных типов волокон обычно незначительны. Небольшие различия (до 5—10 %) могут наблюдаться при сравнении результатов проб, относящихся к центральной и периферической частям мышцы: центральная часть мышцы может содержать больше МС-волокон (De Vries, Housh, 1994).
У одного человека наблюдается относительная однородность структуры различных мышц. Однако наряду со структурной схожестью отдельных мышц конкретного индивидуума могут отмечаться и существенные различия, обусловленные функцией, которую обычно выполняет мышца, и требованием, диктуемым этой функцией мышечным волокнам определенного типа. Так, четырехглавая и икроножная мышцы нижних конечностей, дельтовидная и двуглавая мышца плеча имеют примерно одинаковые соотношения БС- и МС-волокон. В то же время камбаловидная мышца содержит МС-волокон на 25—40 % больше по сравнению с другими мышцами ног, а трехглавая мышца плеча — на 10—30 % больше по сравнению с мышцами пояса верхней конечности (Hanson, 1974). Специальная тренировка приводит к утолщению всех типов волокон, особенно БСб, которые в обычной, жизни малоактивны и очень тяжело вовлекаются в мышечную деятельность (Wilmore, Costill, 2004).
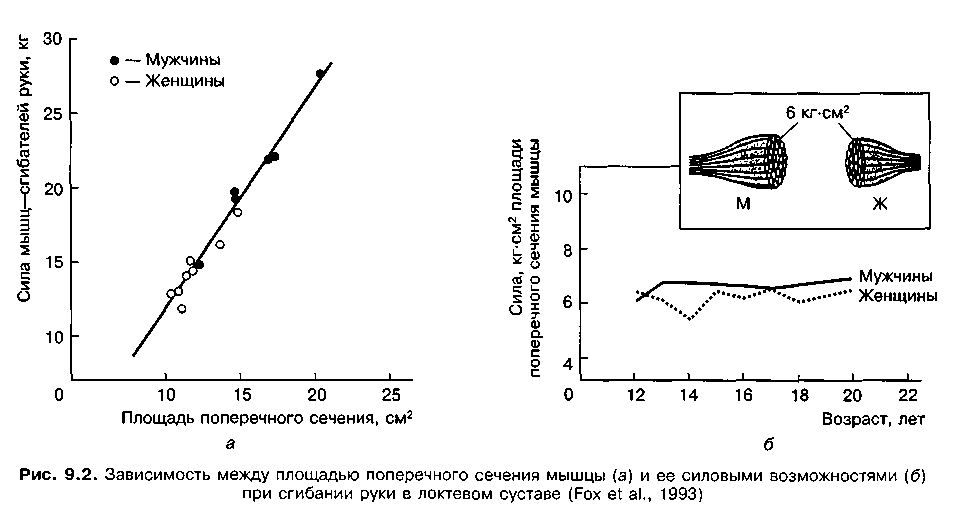
Важно отметить, что структура и возможности мышечных волокон не зависят от пола спортсменов. Зависимость между площадью поперечного сечения и силой является идентичной у мужчин и женщин: на единицу площади поперечного сечения мышцы у мужчин и женщин приходится один и тот же уровень силы (6 кг-см2), в то время как возраст на уровень силовых качеств мышцы существенно не влияет (рис. 9.2).
Спортивная специализация и структура мышечной ткани
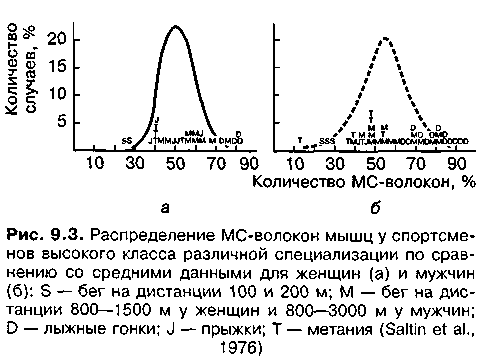
У спортсменов высокого класса наблюдаются различные соотношения мышечных волокон в мышцах, несущих основную нагрузку в данном виде спорта (рис. 9.3—9.5). У бегунов-спринтеров отмечается высокий процент БС-волокон, у лыжников,бегунов на длинные дистанции преобладают МС-волокна, у бегунов на средние дистанции и метателей отмечается относительно равномерное распределение БС- и МС-волокон. Существует строгая зависимость между количеством БС- и МС-волокон в мышечной ткани и спортивными достижениями на спринтерских и стайерских дистанциях. У выдающихся спортсменов не просто преобладают БС- и МС-волокна, а часто наблюдается подавляющее большинство соответствующих волокон. Например, у известного пловца-стайера в дельтовидной мышце оказалось 9 % БС-волокон и 91 % МС-волокон (Counsilman, 1980), среди легкоатлетов отмечались случаи, когда количество БС-волокон в икроножной мышце бегунов-спринтеров достигало 92 %, а у бегунов-стайеров эта мышца на 93—99 % состояла из МС-волокон (Уилмор, Костилл, 2001).
У спортсменов, показывающих высокие результаты в видах спорта, требующих выносливости к длительной работе, отмечается достаточно высокий процент БСа-волокон и незначительный процент БСб-волокон в мышцах, несущих основную нагрузку в тренировочной и соревновательной деятельности. В то же время в мышцах, не несущих нагрузки в данном виде спорта, отмечается нормальное содержание БСб-волокон; в икроножной мышце бегунов на длинные дистанции — 67,1 % МС-волокон; 28,0 % — БСа-волокон и всего 1,9 % — БСб-волокон. В дельтовидной мышце этих же спортсменов было в среднем 68,3 % МС-волокон; 14,3 % — БСа-волокон и 17,4% — БСб-волокон (Jansson, Kaiser, 1977). 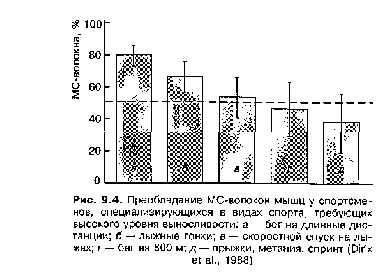
Это является достаточно веским основанием для предположения, что исчезновение БСб-волокон — часть адаптивной реакции организма при тренировке выносливости.

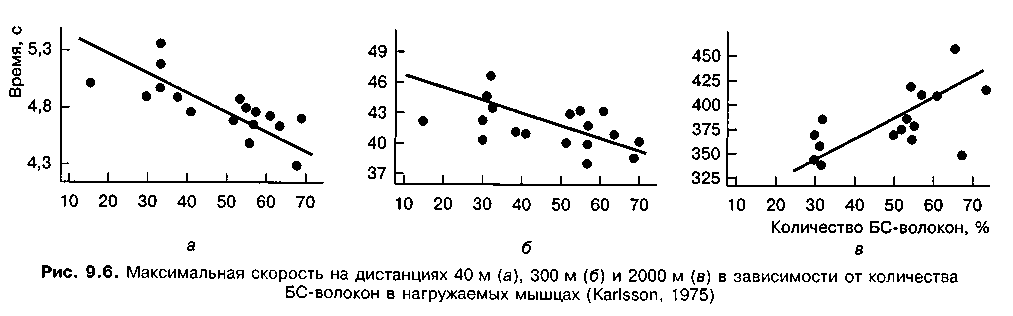
Существует тесная связь между уровнем скоростных способностей спортсменов и количеством БС-волокон в нагружаемых мышцах (рис. 9.6). Так, между скоростью бега на дистанции и количеством БС-волокон существует тесная корреляционная связь (г = 0,73). Увеличение длины дистанции связано со снижением этой связи (г = 0,45). При увеличении дистанции до 2000 м, время пробегания которой у испытуемых превышало 5 мин, связь приобретает отрицательный характер: наличие большого количества БС-волокон в нагружаемых мышцах отрицательно влияет на результат. Таким образом, количество мышечных волокон определенного типа в значительной мере обусловливает достижения спортсменов в различных видах соревнований. В спринтерской работе скоростно-силового характера (бег на 100 м, бег на коньках на 500 м, плавание на 50 м, легкоатлетические прыжки и др.) большое значение имеют БСб-волокна. В беге на 400 и 800 м, плавании на 100 и 200 м и др. очень велика роль БСа-волокон, функциональные свойства которых отвечают требованиям эффективной соревновательной деятельности в этих видах соревнований. Успех в стайерских дисциплинах разных видов спорта в решающей мере определяется количеством МС-волокон.
Структура мышечной ткани во многом зависит от квалификации спортсменов. Например, у тяжелоатлетов различной квалификации отмечается разный процент БС-волокон. У спортсменов низкой квалификации таких волокон обычно не более 45—55 %. Спортсмены международного класса имеют более высокий процент волокон — 60—70 % (Foucart et al., 1984).
 Изменения в мышечных волокнах под влиянием нагрузок различной величины и направленности
Изменения в мышечных волокнах под влиянием нагрузок различной величины и направленности
В настоящее время вопрос о превращении одного типа мышечных волокон в другой под влиянием специфической тренировки остается до конца не решенным. Специалисты склоняются к мнению, что соотношение мышечных волокон различного типа у человека обусловлено генетически. Что касается влияния интенсивной тренировки определенной направленности (развитие выносливости к длительной работе, скоростно-силовой), то она приводит к существенному изменению морфологических, физиологических и биохимических свойств мышечных волокон. Под влиянием тренировки, направленной на повышение выносливости, трансформация свойств мышечных волокон различных типов происходит в следующем порядке: БСб-волокна приобретают свойства БСа-волокон, а БСа-волокна — свойства МС-волокон. Силовая подготовка вызывает обратный процесс: МС-волокна приобретают свойства БСа-волокон, а БСа-волокна — соответственно свойства БСб-волокон.
Оба типа мышечных волокон имеют характеристики, которые могут быть изменены в процессе тренировки. Размеры и объем БС-волокон увеличиваются под влиянием тренировки скоростного, скоростно-силового и силового типа, в результате чего их процентное соотношение в площади поперечного сечения мышцы возрастает (Zesper et al., 2000). Одновременно повышается их гликолитическая способность. При тренировке на выносливость оксидативный потенциал МС-волокон может возрастать в 2—4 раза.
Среднее количество капилляров вокруг МС-и БСа-волокон составляет 4, а вокруг БСб-воло-кон — 3. У спортсменов высокого класса, выступающих на длинных дистанциях, в дельтовидных мышцах пловцов и икроножных мышцах бегунов были обнаружены МС-волокна, каждое из которых снабжалось 5—6 капиллярами (Nygaard, Goricke, 1976). Эффект напряженной тренировки аэробного и смешанного (аэробно-анаэробного) характера проявляется в увеличении количества капилляров на мышечное волокно или на квадратный миллиметр мышечной ткани. Здесь выявляются два механизма: увеличение количества капилляров; если же возможности этого механизма исчерпаны или невелики, то происходит уменьшение размера мышечных волокон (Hoppeler et al., 1990).
Длительная и напряженная тренировка аэробной направленности приводит к изменению соотношения волокон различных типов. Об этом косвенно свидетельствуют результаты многочисленных исследований композиции мышечной ткани, несущей основную нагрузку в тренировочной и соревновательной деятельности в сравнении с тканью, не подвергавшейся активной тренировке. У гребцов на байдарках и пловцов-стайеров в дельтовидной мышце регистрировалось до 60— 70 % МС-волокон, а в широких мышцах бедра таких волокон было не более 45—60 %. У велоси-педистов-шоссейников, лыжников, бегунов-стайеров картина противоположная: в икроножной мышце регистрировалось до 60—80 % и более МС-волокон, а в дельтовидной мышце и трехглавой мышце плеча их количество у этих же спортсменов не превышало 50—60 %.
МС-волокна очень слабо подвержены скоростной тренировке. Так, спортсмены, в мышцах которых содержится малое количество БС-волокон, слабо приспосабливаются к скоростной работе даже после напряженной тренировки скоростного характера. Например, высота прыжка вверх у таких спортсменов, специализирующихся в плавании, обычно не превышает 45—50 см, в то время как у спортсменов с большим количеством БСа- и БСб-волокон она редко бывает ниже 70 см (Counsilman, 1980).
Проведены интересные исследования, отражающие структуру, функциональные возможности и особенности адаптации мышечных волокон различного типа (Schon et al., 1978). Анализ структуры и распределения митохондрий и частиц нейтральных жиров с помощью электронной микроскопии показал, что в БСб-волокнах отмечается значительно меньшее количество митохондрий по сравнению с БСа-волокнами и, особенно, МС-во-локнами. Частицы жиров в БСб-волокнах отсутствуют вообще, в БСа-волокнах их немного, а в МС-волокнах они встречаются в очень большом количестве. У хорошо тренированных бегунов на длинные дистанции по сравнению со студентами спортивного вуза в БСб-волокнах заметных различий указанных выше компонентов не отмечено. В БСа-волокнах у бегунов большее содержание митохондрий и частиц нейтральных жиров, в МС-волокнах доля центральных митохондрий — в 2 раза, а периферийных — в 3 раза выше, чем у студентов спортивных вузов. У бегунов выявлено увеличение количества и объема митохондрий, а также в 3 раза более высокое содержание частиц нейтральных жиров. У них также установлены положительные корреляции массы митохондрий с величинами максимального потребления кислорода и отрицательные корреляции — с величиной лактата при субмаксимальной нагрузке. Все это свидетельствует о значительном воздействии характера тренировочной и соревновательной деятельности на характеристики мышечных волокон, существенно влияющие на их функциональные возможности.
Рассматривая гипертрофию мышечных волокон в качестве одного из основных путей адаптации мышц, следует указать, что гипертрофия МС-волокон, связанная, прежде всего, с увеличением размеров миофибрилл, возрастанием количества и плотности митохондрий, приводит к увеличению удельного веса в мышечной массе МС-волокон и, как следствие, к повышению выносливости и уменьшению скоростных способностей мышц. С другой стороны, гипертрофия БС-волокон приводит к увеличению их удельного веса в мышце по сравнению с МС-волокнами и способствует повышению ее скоростного потенциала (De Vries, Housh, 1994). При этом характер нагрузки определяет, какие из мышечных волокон претерпевают значительные изменения. Продолжительные нагрузки относительно невысокой интенсивности преимущественно приводят к увеличению объемной плотности митохондрий МС- и БСа-волокон. Интенсивная интервальная работа в основном способствует возникновению изменений в БСб-волок-нах (Dudley et al., 1982; Виру и др., 1993).
Гипертрофия мышцы связана с рядом изменений, в числе которых прежде всего следует отметить увеличение резервов актиновых и, особенно, миозиновых нитей, увеличение количества миофибрилл и кровеносных капилляров в волокне.
Длительное время считалось, что количество мышечных волокон в каждой мышце детерминировано генетически и остается неизменным в течение всей жизни (Gollnick et al., 1981; Timson et al., 1985). Однако в отдельных работах (Гудзь, 1975, 1977) была продемонстрирована возможность гиперплазии мышц в ответ на большие физические нагрузки. В дальнейшем появилось достаточное количество доказательств того, что напряженная и длительная силовая тренировка приводит не только к гипертрофии мышечных волокон, но и к увеличению их количества. Силовая тренировка с большими отягощениями и небольшим количеством повторений в течение двух лет не только привела к увеличению мышечной силы и гипертрофии мышц, но и к достоверному увеличению (на 9 %) общего количества мышечных волокон (Gonyea et al., 1986). Возможность процесса гиперплазии у людей была показана в исследованиях с участием культуристов (Larsson, Tesch, 1986; Tesch, 1991).
Однако вопрос о возможности гиперплазии мышечных волокон под влиянием тренировки все же не является до конца решенным. Действительно, при гипертрофии мышечные волокна могут расщепляться, но есть доказательства того, что расщепление не касается всей длины мышечного волокна и не может являться основанием для утверждения, что имеет место процесс гиперплазии, т. е. деления и увеличения общего количества мышечных волокон. Это дает основание предположить, что количество мышечных волокон у человека является генетически детерминированным и в результате тренировки не изменяется. Косвенным подтверждением этого является и факт, согласно которому увеличение площади поперечного сечения мышечных волокон происходит строго пропорционально увеличению площади поперечного сечения всей мышцы (Мак-Комас, 2001).
Принципиально важным вопросом для спортивной практики является возможность трансформации мышечного фенотипа, преобразования волокон одного типа в волокна другого. Структура и функциональные возможности мышечных волокон различного типа обусловливаются особенностями их нервной импульсации, которая и определяет, будет ли данное волокно иметь свойства быстросокраща-ющегося или медленносокращающегося. Если БС-волокна стимулируются по принципу импульсации МС, то в них повышается активность оксидативных ферментов. И, наоборот, стимуляция МС-волокон по принципу БС приводит к повышению активности гликолитических ферментов (Pette, 1984).
Исследования на животных показали, что иннервация БС-волокон путем переноса в нерв (с помощью специальных электродов) электроимпульсов с частотной характеристикой, соответствующей иннервации МС-волокон, приводит к изменению структурных и функциональных свойств волокон (Brown et al., 1983). В волокнах увеличивается плотность капилляров (Hudlicka et al., 1980), повышается содержание миоглобина (Pette, 1984), что приводит к изменению цвета бледных волокон, которые становятся ярко-красными. Оксидативные способности волокон повышаются за счет увеличения активности ферментов, окисляющих субстраты. Одновременно угнетается анаэробная способность волокон в связи со снижением активности ферментов, участвующих в процессе гликолиза (Buchegger et al., 1984).
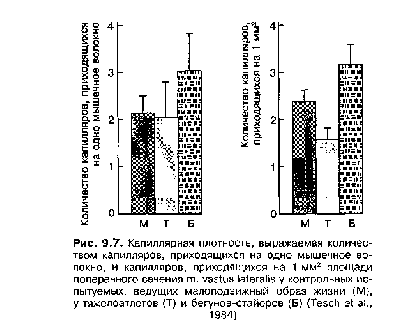
Адаптация различных типов мышечных волокон находится в строгой зависимости от направленности тренировочного процесса. Например, у велоси-педистов-шоссейников не только отмечается большой процент МС-волокон по сравнению с лицами, не занимающимися спортом, или спортсменами других специализаций, но и существенная гипертрофия (до 20—30 %) этих волокон при одновременной атрофии или перестройке БСб- и БСа-волокон. В результате при среднем проценте МС-волокон у велосипедистов-шоссейников около 70— 75 % они могут занимать до 85—90 % и более общего поперечного сечения мышц, несущих основную нагрузку при педалировании. Увеличение объема МС-волокон в результате тренировки стайерского типа сопровождается и рядом других изменений, происходящих в мышечной ткани. Одним из важнейших является значительное увеличение количества капилляров в работающих мышцах (рис. 9.7).
Важно отметить, что рост новых капилляров в результате работы, направленной на повышение выносливости, отмечается не только в МС-во-локнах, но и БСа- и, даже, БСб-волокнах. Правда, изменения в БСб-волокнах выражены в значительно меньшей мере, чем в МС- и БСа-волокнах. В результате резко возрастает емкость капиллярного ложе, что ускоряет процесс доставки кислорода и питательных веществ к мышечным волокнам и выведения остаточных продуктов метаболизма.
Основным эффектом увеличения капилляриза-ции мышц является замедление кровотока через капилляры, что способствует улучшению передачи кислорода в мышечные волокна, увеличению арте-риовенозной разницы.
В результате интенсивной и продолжительной тренировки, направленной на развитие выносливости, изменения активности аэробного и анаэробного путей обмена качественно напоминают метаболические трансформации, отмеченные в результате искусственно стимулированных мышц. Значительные изменения отмечаются в митохондриях, плотности капиллярной сети, в составе миозина (Green et al., 1983). Однако адаптационные перестройки выражены гораздо меньше, чем в тех случаях, когда обеспечивается постоянная стимуляция (Howald, 1982; Pette, 1984). В связи с этим существует мнение (Henriksson, 1992), что окислительные способности тренированных мышц спортсменов высокого класса, специализирующихся в видах спорта, требующих высокого уровня аэробных возможностей, составляют лишь 50— 70 % теоретически достижимого уровня.
Известно, что БС-волокна используют в единицу времени намного больше энергии, чем МС-волокна. Воздействие специальной тренировки, проявляющееся в трансформации БС-волокон в МС-волокна, представляет собой определенный вид экономизации функций, так как создает условия для выполнения продолжительной работы с меньшими затратами энергии (Streter et al., 1982). Однако эта экономизация связана с существенным уменьшением скорости сокращений (Pette, 1984; Roberts, Robergs, 2002).
Таким образом, тренировка на выносливость способна значительно повысить возможности окислительного способа энергообеспечения не только БСа-, но и БСб-волокон. Более того, тренированные на выносливость БСа-волокна по своим окислительным способностям могут даже превышать показатели МС-волокон, характерные для нетренированного человека (Essen et al., 1975; Jansson, Kaiser, 1977). Большие объемы работы на развитие выносливости могут даже привести к такой трансформации БСб-волокон, что их вообще не удастся обнаружить в поперечном срезе мышцы. Естественно, что эти изменения приводят к резкому снижению скоростных возможностей мышц. Специалисты считают, что восстановление мышечных БС-волокон в принципе возможно, однако очень сложно и в настоящее время неизвестно, какие средства являются для этого наиболее эффективными (Noth, 1992). Одной из основных проблем трансформации быстросокращающейся мышечной ткани в медленносокращающуюся является то, что БС-волокна вследствие высокого порога возбуждения значительно реже и сложнее включаются как в повседневную, так и в специальную тренировочную и соревновательную мышечную деятельность (Dum et al., 1985; Platonov, Bulatova, 2003).
Вместе с тем никакой специальной тренировкой, связанной с развитием выносливости, невозможно добиться в БС-волокнах таких изменений, которые характерны для хорошо тренированных МС-волокон, и при прочих равных условиях спортсмены с большим количеством МС-волокон всегда будут иметь преимущество на длинных дистанциях над спортсменами, у которых таких волокон значительно меньше.
В зависимости от типа мышечных волокон их гипертрофия под влиянием силовой тренировки носит избирательный характер. Силовая тренировка, направленная на повышение мышечной силы за счет гипертрофии мышечной ткани, приводит к увеличению поперечного сечения всех типов мышечных волокон. Однако наиболее интенсивно этот процесс происходит в БС-волокнах: в результате б-месячной тренировки поперечное сечение быстросокращающихся мышечных волокон обеих групп (БСа и БСб) может увеличиться на 20—50 %. Такой широкий диапазон различий зависит от исходного уровня развития мышц, индивидуальных особенностей занимающихся, эффективности системы тренировки и др. За этот же период поперечное сечение медленно сокращающихся мышечных волокон может быть увеличено только на 5—10 %. Для спортивной практики важным является вопрос о времени, в течение которого происходит морфо-функциональная перестройка мышечных волокон различного типа под влиянием напряженной специфической тренировки. Первые серьезные изменения отмечаются уже на второй неделе, а после месяца тренировки проявляется весь комплекс адаптационных реакций морфологического, функционального и биохимического характера.
Координация деятельности двигательных единиц — важный механизм адаптации мышц к физическим нагрузкам
Эффективная работа, направленная на совершенствование функциональных возможностей мышц, диктуемая требованиями тренировочной и соревновательной деятельности в различных видах спорта, может быть осуществлена лишь на основе четкого понимания механизмов нервной регуляции их деятельности. Усиление активности работы мышц связано с рекрутированием необходимого объема двигательных единиц и усилением стимуляции уже работающих мышечных волокон.
В настоящее время можно считать общепризнанной теорию последовательного рекрутирования двигательных единиц, предусматривающую первоочередное вовлечение в работу мелких мотонейронов и соответственно двигательных единиц. С увеличением интенсивности работы она обеспечивается вовлечением более крупных двигательных единиц.
Между мелкими и крупными двигательными единицами существует большая разница. Например, самая крупная двигательная единица в икроножной мышце человека способна развить напряжение в 200 раз большее, чем самая мелкая. Для общего усиления работы мышц активизируются более крупные двигательные единицы, что обеспечивает большой прирост напряжения, т. е. по мере увеличения общего напряжения оно обеспечивается меньшим числом дополнительных единиц. Разумеется, при предельных или околопредельных напряжениях двигательные единицы вовлекаются не последовательно, а активизируются практически одновременно (Эвартс, 1984, Behnke, 2001).
Результаты исследований свидетельствуют о том, что фактором, определяющим количество и тип необходимых для использования волокон, является величина сопротивления. Нервная система регулирует деятельность мышц в зависимости от того, какую силу должна развить мышца, а не от скорости ее сокращения. Объяснить это можно тем, что МС-волокна способны перемещать биозвенья с очень большой скоростью (более 1000° в 1 с), но только при условии незначительных силовых проявлений (Costill et al., 1980; De Vries, Housh, 1994).
Таким образом, первыми в работу вовлекаются двигательные единицы медленного сокращения — самые мелкие из типов двигательных единиц. Если они не способны развить необходимую силу, рекрутируются двигательные единицы быстрого сокращения (рис. 9.8): например, во время медленного плавания большая часть продвига-
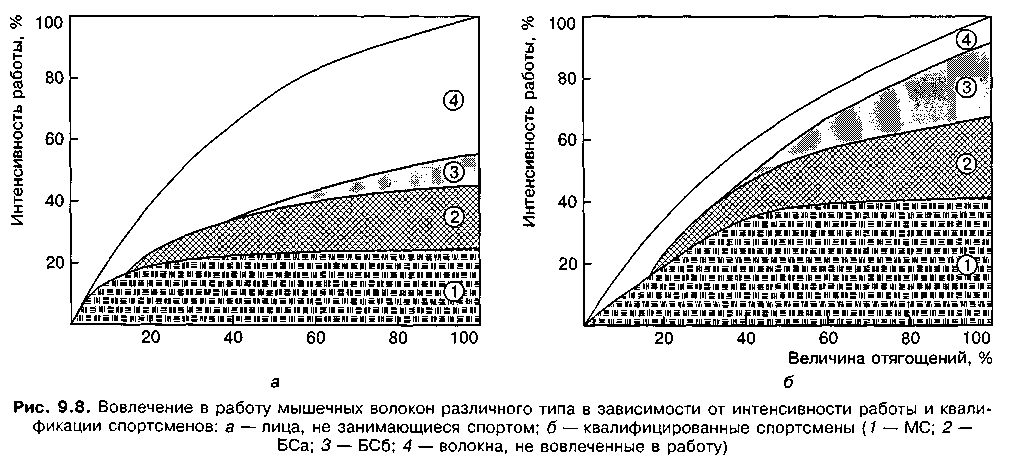
ющей силы создается МС-волокнами. С ростом скорости и уровня приложения усилий в работу вовлекаются БСа-волокна. Когда необходимо проявление максимальной силы, в работе участвуют, кроме первых двух типов, БСб-волокна. Педалирование на велоэргометре с высокой скоростью, но без отягощения осуществляется преимущественно за счет МС-волокон; педалирование при большом отягощении вовлекает в работу все три типа волокон как при низкой, так и при высокой скорости (Platonov, Bulatova, 2003).
Разница величин порога возбуждения мышечных волокон различных типов предопределяет последовательность их вовлечения в активную мышечную деятельность. Небольшие проявления силы могут быть обеспечены двигательными единицами, имеющими низкий порог возбуждения. Для увеличения проявлений силы или длительного поддержания заданного мышечного напряжения требуется включение двигательных единиц с более высоким порогом возбуждения (Thomas et al.,
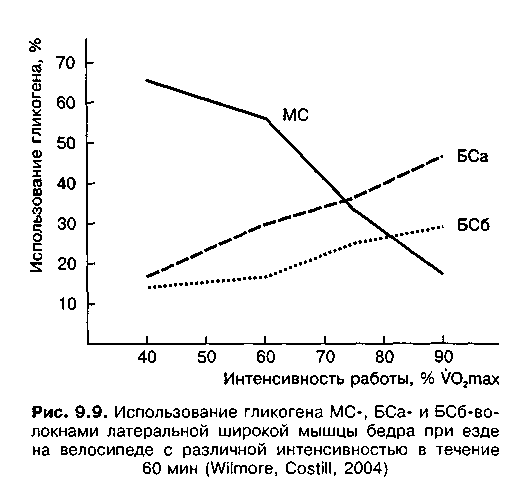
1987; Kernell, 1990). О зависимости рекрутирования различных типов мышечных волокон от интенсивности работы свидетельствуют данные о расходе мышечного гликогена в мышечных волокнах. При езде на велосипеде с относительно невысокой интенсивностью (40—60 % V02max) наиболее активными были МС-волокна. При работе с высокой интенсивностью (90 % V02max) основную нагрузку несли БСа-волокна (рис. 9.9).
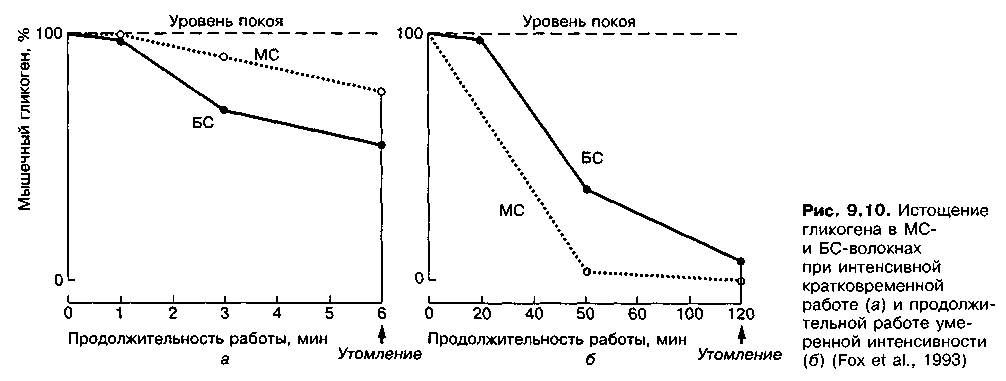
При интенсивной работе смешанного анаэробно-аэробного характера (интенсивный бег в течение б мин до явного утомления) преимущественно истощаются запасы гликогена в БС-волокнах. Когда же выполняется продолжительная работа аэробной направленности (2-часовой бег до явного утомления), полностью исчерпываются запасы мышечного гликогена в МС-волокнах (рис. 9.10).
Принято считать, что МС- и БСа-волокна максимально вовлекаются в работу, когда ее интенсивность достигает 80—85 % V02max (Saltin, Gollnick, 1983). Для максимальной активации БСб-волокон требуется предельная интенсивность работы (Henriksson, 1992; Wilmore, Costill, 2004). Однако из этого общего принципа имеются исключения. При очень быстрых и интенсивных рефлекторных или произвольных мышечных действиях может наблюдаться обратный порядок: двигательные единицы с высоким порогом возбуждения могут активизироваться без активизации небольших единиц с низким порогом возбуждения. Более того, может наблюдаться подавление активности двигательных единиц с низким порогом возбуждения (Mellah et al., 1990).'
От одного двигательного нейрона могут иннер-вироваться несколько сотен мышечных волокон, одновременно мышечные волокна обычно имеют окончания как собственного, так и смежных нейронов. Импульс, поступающий к волокнам, составляющим данную двигательную единицу, охватывает и рядом расположенные двигательные единицы, число которых в крупных мышцах может достигать
нескольких тысяч (Эвартс, 1984). Способность человека дифференцировать интенсивность мышечного сокращения путем включения минимально необходимого количества двигательных единиц находится в числе важнейших реакций адаптации мышц и в значительной мере обусловливает эффективность внутримышечной координации.
Продолжительная мышечная работа в случае эффективной адаптации связана с попеременным включением различных двигательных единиц. При усталости отдельных из них их функции выполняют другие, если характер работы допускает такую компенсацию, а при снижении возможностей всех двигательных единиц, участвующих в выполнении конкретной работы, поддержание работоспособности связано с усилением нервной импульсации (Зимкин, 1984). Развитие утомления и истощение запасов гликогена в МС-волокнах в процессе длительной работы невысокой интенсивности требуют для продолжения упражнения интенсивного вовлечения БСа-волокон, а в его заключительной части и БСб-волокон (Wilmore, Costill, 2004).
Не менее важной для эффективной тренировочной и соревновательной деятельности в различных видах спорта является реакция адаптации, связанная с увеличением способности ЦНС к мобилизации двигательных единиц в мышцах. При выполнении упражнений с околопредельными или предельными отягощениями удается вовлечь в работу максимально возможное количество двигательных единиц. Обусловливается это прежде всего тем, что при произвольных сокращениях проявления силы зависят от состава мышечных волокон, вовлеченных в работу, и частоты активизации двигательных единиц: чем большее количество мышечных волокон различных типов вовлечено в работу и чем интенсивней их нервная импульса-ция, тем выше будут показатели развиваемой силы (Alway et al., 1990; Moritani, 1992).
Активация мышечных БС-волокон происходит не только при выполнении работы высокой интенсивности, но и при работе относительно невысокой интенсивности, когда истощаются запасы гликогена в мышечных МС-волокнах (Мак-Комас, 2001).
Эффект долговременной адаптации к физическим нагрузкам силового характера проявляется в резком увеличении количества двигательных единиц, вовлекаемых в работу. Так, у нетренированного человека число двигательных единиц, которые могут быть мобилизованы при максимальных силовых напряжениях, обычно не превышает 25—30 %, а у хорошо тренированных к силовым нагрузкам лиц процент вовлеченных в работу моторных единиц может превышать 80—90 %. В основе этого явления лежит адаптация центральной нервной системы, приводящая к повышению способности моторных центров мобилизовывать большее число мотонейронов и к совершенствова-' нию межмышечной координации (Зимкин, 1984; Hoffman, 2002).
Другим направлением адаптации мышц является улучшение межмышечной координации, связанное с совершенствованием деятельности мышц-агонистов, обеспечивающих выполнение движения; мышц-синергистов, способствующих выполнению движения, и мышц-антагонистов, препятствующих выполнению движения. Рациональная координация деятельности этих групп мышц не только обеспечивает высокую силу и скорость сокращения, точность выполнения движения, но и обусловливает экономичность работы.
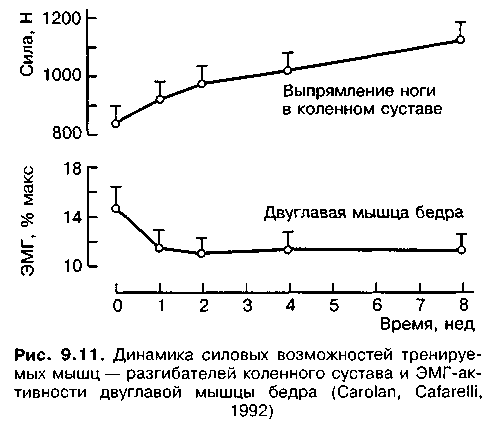
Экономичность работы в той ее части, где это связано с деятельностью мышц-антагонистов, зависит от эластичности мышц, подвижности в соответствующих суставах. Так, незначительная эластичность мышц-антагонистов значительно затормаживает маховые движения в конце фазы, в силу чего снижается их амплитуда и экономичность (Gambetta, 1987). Систематическая тренировка приводит к устранению излишнего напряжения мышц-антагонистов при выполнении различных упражнений и одновременно обеспечивает эффективную координацию деятельности мышц-аго-нистов и мышц-синергистов в достижении конечного заданного эффекта (Алтер, 2001). Например, под влиянием силовой тренировки увеличивается частота разрядки мотонейронов, возрастает стабильность импульсации двигательных единиц с высоким порогом возбудимости, улучшается синхронизация деятельности мышц-синергистов, снижается реактивность мышц-антагонистов. На рис. 9.11 видно, как под влиянием силовой тренировки увеличение силы тренируемых разгибателей коленного сустава сопровождается уменьшением активности двуглавой мышцы бедра, препятствующей разгибанию коленного сустава.
Принято считать, что максимальная активизация деятельности мышц-агонистов и мышц-синер-гистов при минимальной активности мышц-антагонистов является основной для эффективной двигательной деятельности в спорте. Однако это справедливо далеко не для всех двигательных действий. Когда речь идет о работе с относительно невысокой интенсивностью или выполнении движений с максимальной амплитудой, то минимизация активности мышц-антагонистов — важный фактор в обеспечении результативности двигательной деятельности. В тех же условиях, когда требуется максимальное усилие (например, в тяжелой атлетике различных видах борьбы) совместное возбуждение, при котором активизируются и мышцы-антагонисты оказывается более целесообразным, так как обеспечивает значительно большую жесткость и устойчивость сустава, более высокие показатели проявления силы и уровня ста-тодинамической устойчивости (Энока, 2000).
Связь эффективности внутримышечной и межмышечной координации с экономичностью работы и уровнем мастерства можно также проследить по показателям активности мышц спортсменов различной квалификации при выполнении стандартных нагрузок: у спортсменов высокой квалификации отмечается значительно меньшая электрическая активность мышц по сравнению с менее квалифицированными спортсменами. В то же время при выполнении предельных нагрузок у спортсменов высокого класса выявляется значительно более высокая активность мышц-агонистов и мышц-синер-гистов по сравнению с лицами, не занимающимися спортом, или спортсменами невысокой квалификации. В отношении мышц-антагонистов проявляется обратная зависимость: чем выше квалификация спортсмена, тем меньше активность мышц.
В результате суммарной перестройки морфологических, биохимических и физиологических механизмов, обусловливающих эффективность приспособления организма человека к работе силового характера, сила мышц может возрастать в 2—4 раза (Зимкин, 1984). Адаптация организма к силовой тренировке обусловлена изменениями в мышцах, нервной системе, костной ткани. Увеличение силы связано как с гипертрофией мышц, так и с увеличением плотности сокращающихся элементов внутри клетки, изменением соотношения актина и миозина (Penman, 1970). Морфологические и функциональные изменения в нервной системе в первую очередь сводятся к разветвлению мотонейронов, увеличению ганглиозных клеток. Изменения костной системы связаны с увеличением плотности костей, их эластичности, гипертрофией костных выступов в местах прикрепления сухожилий мышц. Эти изменения особенно ярко проявляются у представителей скоростно-силовых видов спорта: тяжелоатлетов, метателей, бегунов-спринтеров. В то же время у пловцов существенных изменений костей по сравнению с лицами, не занимающимися спортом, не отмечается (Westlin, 1970).
В результате специальной тренировки сила увеличивается в 1,5—2,5 раза по сравнению с мышечной массой (Hollmann, Hettinger, 1980). Существует два относительно самостоятельных механизма повышения силы. Первый механизм связан с морфофункциональными изменениями в мышечной ткани — гипертрофией и в определенной мере гиперплазией мышечных волокон; второй предусматривает совершенствование способностей нервной системы синхронизировать возможно большее количество двигательных единиц, что приводит к увеличению силы без увеличения объема мышц. Гипертрофия обусловливается постоянным чередованием в силовой тренировке процессов белкового расщепления и белкового синтеза с преобладанием последнего. Изменения нервной регуляции проявляются в улучшении синхронизации деятельности двигательных единиц, изменении последовательности их рекрутирования, увеличении максимального уровня интегрированной электромиограммы. Эти нейрогенные реакции адаптации обеспечивают существенное увеличение максимальной силы при незначительном увеличении мышечной массы (Sale, 1992).
На различных этапах силовой подготовки увеличение силы происходит за счет преимущественного использования либо нервных, либо мышечных механизмов адаптации. На первом этапе прирост силы связан, прежде всего, с повышением возможности нервной стимуляции мышечной деятельности. В дальнейшем постепенно разворачивается процесс мышечной гипертрофии.
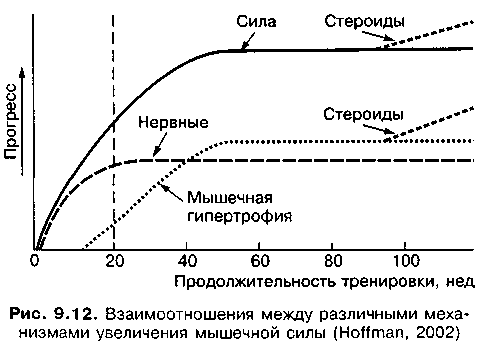
После исчерпания возможностей обоих путей увеличения силы спортсмены часто прибегают к использованию анаболических стероидов, что интенсифицирует прирост мышечной силы (рис. 9.12).
Следует учитывать, что разные типы мышечных волокон имеют различный порог раздраже ния. Минимальный порог раздражения (10—15 Гц) имеют МС-волокна, обеспечивающие работу на уровне 20—25 % максимальной статической силы. Развитие 60 % максимальной статической силы вовлекает в работу БС-волокна и связано с частотой 20—45 Гц, а предельные силовые проявления, требующие вовлечения в работу всего мышечного массива, требуют частоты 45—60 Гц. Порог раздражения зависит и от объема двигательных единиц: чем меньше двигательная единица, тем ниже порог ее раздражения (Green et al., 1983). Именно поэтому упражнения с отягощениями, составляющими 20—30 % уровня максимальной силы, не только не способствуют развитию этого качества, но и по отношению к спортсменам с высоким уровнем силовых качеств даже не позволяют сохранить им ранее достигнутый уровень. Критический порог напряжения, обеспечивающий повышение максимальной силы для нетренированных лиц, лежит в пределах 50—60 % максимальных возможностей в соответствующем упражнении. Вполне естественно, что у спортсменов высокого класса критический порог существенно возрастает и достигает 80—90 % (Platonov, Bulatova, 2003).
Гипертрофия различных типов мышечных волокон определяется методикой тренировки. Микроскопическому исследованию были подвергнуты мышцы культуристов, которые в тренировке использовали небольшие отягощения при большом количестве повторений и небольшой скорости движений. Обнаружилось, что МС-волокна были гипертрофированы, в то время как БС-волокна не увеличили своего объема. Применение больших отягощений при небольшом количестве повторений и высокой скорости движений, наоборот, приводит к избирательной гипертрофии БС-волокон, а объем МС-волокон остается без существенных изменений (Counsilman, 1980; Tesch, 1991).
Долговременная адаптация мышц при предельных и околопредельных скоростно-силовых нагрузках, приводящих к развитию силы, связана со значительной гипертрофией мышц, особенно БС-волокон, что приводит к существенному увеличению их площади в поперечном срезе мышечной ткани (Tesch, Karlsson, 1984). При таких нагрузках не отмечается заметных изменений васкуляриза-ции мышц, не изменяется мощность системы митохондрий в мышцах. Одновременно происходит перестройка энергетического метаболизма мышечных волокон в направлении увеличения мощности системы гликолитического ресинтеза (Hollmann, Hettinger, 1980; Wilmore, Costill, 2004).
Гипертрофии БС-волокон способствуют различные упражнения: с дополнительными отягощениями или выполняемые с использованием специальных тренажеров, целостные действия в борьбе, удары в футболе, броски в гандболе и водном поло, метание молота, толкание ядра, спринтерский бег, старт в плавании.
Важно учитывать, что сила, приобретенная с помощью упражнений при высоких скоростях движений, имеет перенос на более низкие скорости (Counsilman, 1980), тогда как сила, развитая использованием упражнений при низких скоростях движения, переноса на двигательные действия, выполняемые с высокой скоростью, не имеет (Pipes, Wilmore, 1976; Платонов, Булатова, 1995). Одновременно при тренировке с высокими скоростями движений отмечается большее снижение жировой ткани по сравнению с тренировкой на низких скоростях.
Специальными исследованиями установлено, что прирост силовых качеств в течение первых дней тренировки связан с совершенствованием внутри- и межмышечной координации, что обусловлено привлечением к работе большего количества двигательных единиц, оптимизацией работы мышц-синергистов, устранением иннервации антагонистов. При этом из числа синергистов основную нагрузку несут мышцы, которые в состоянии обеспечить наиболее эффективное выполнение данного движения с учетом его направленности (Кузнецов, 1970; Энока, 2000).
Адаптация костной
и соединительной тканей
Интенсивное исследование адаптационных перестроек костной и соединительной тканей под влиянием различных факторов стало проводиться лишь в последние десятилетия. До этого считалось, что костная и, особенно, волокнистая соединительная ткань не предрасположены к адаптации. Однако в настоящее время установлено, что кости, сухожилия, связки весьма чувствительны к механическим нагрузкам и реагируют на них соответствующими структурными и функциональными изменениями.
В структуре костной ткани выделяют минеральные, органические и жидкостные компоненты. Минералы (кристаллы кальция гидроксиапатита) составляют около 50 % общего объема кости и обеспечивают ее твердость. Органические элементы составляют 40 % объема кости (преимущественно коллаген — 95 %) и обеспечивают ее эластичность. Оставшиеся 10 % приходятся на сосудистые каналы и клеточные пространства.
Минеральное содержание отличает кость от других соединительных тканей, в частности связок и сухожилий, которые представляют собой плотные волокнистые ткани, состоящие в основном из коллагена.
Сухожилия — белые коллагеновые полоски, соединяющие мышцы с костями, через которые передаются силы мышечного сокращения в костную систему. Важной является способность сухожилий обеспечивать проявление дополнительной силы за счет их эластичных свойств, проявляемых в случаях околопредельного или предельного предварительного растяжения мышц (Stone, 1992). Связь сухожилия с костью происходит путем постепенного перехода от сухожилия к волокнистому хрящу, затем к минерализованному хрящу и кости. Коллагеновые волокна сухожилия могут также непосредственно смешиваться с коллагеновыми волокнами надкостницы. Присоединение сухожилия к мышце осуществляется посредством синапса, представляющего собой многослойное сложное соединение, образованное филаментом актина конечного саркомера и коллагеновыми волокнами сухожилия (Ovalle, 1987). Интересно, что МС-во-локна мышц имеют значительно большую синапсо-вую (соединительную) поверхность по сравнению с БС-волокнами (Tidball, Daniel, 1986).
Связки, представляющие собой пучки коллаге-новых волокон, соединяют соседние кости и могут быть внешними и внутренними относительно суставной капсулы.
В настоящее время нельзя с достоверностью говорить о механизмах, определяющих способность костей и волокнистой соединительной ткани перестраиваться под влиянием внешних и внутренних факторов. Однако наличие таких механизмов и их результатов в виде существенных изменений в структуре и функциях костей, связок и сухожилий несомненно.
Адаптацию костной и волокнистой соединительной ткани могут обусловливать факторы — питательные, гормональные и функциональные. Например, минеральные вещества и витамины способствуют осаждению минеральных веществ в костях, повышают их плотность (Stone, 1992). Большое влияние на состав соединительной ткани оказывают анаболические стероиды (Wood et al., 1988). Однако основное влияние на адаптацию костной и волокнистой соединительной тканей оказывают физические нагрузки определенного характера и направленности.
Физические нагрузки являются основным фактором, определяющим увеличение костной массы у людей. Степень адаптации костной ткани находится, естественно, в генетически обусловленных пределах, в прямой зависимости от величины нагрузок (Lanyon, 1987). Среди компонентов нагрузки, способствующих увеличению плотности минералов кости, основным является величина отягощения. Экспериментально установлено (Конрой и др., 1996), что адаптация костной массы юных штангистов на 30—50 % (в зависимости от анатомического участка и индивидуальных особенностей спортсмена) зависит от силы, развиваемой при выполнении упражнений. Оставшуюся долю можно объяснить множеством факторов, начиная от генетических особенностей и заканчивая неспособностью применявшихся силовых упражнений эффективно влиять на определенные участки кости.
Адаптационные изменения в костной ткани под влиянием физических нагрузок чаще всего связаны с повышением ее прочностных свойств. Наиболее важные изменения сводятся к увеличению размеров, внешней формы и внутренней структуры компактного и губчатого вещества костей (Солодков, Судзиловский, 1996), плотности минералов (Wiliams et al., 1984). Обнаружены различия в минеральном составе, плотности и массе костей доминирующих конечностей по сравнению с недоминирующими, а наибольшие изменения отмечаются в тех участках скелета, которые подвергаются наиболее интенсивным механическим воздействиям (Montoye et al., 1980).
Плотность костей в значительной мере определяется квалификацией спортсменов, спецификой тренировочной и соревновательной деятельности в различных видах спорта. У спортсменов высокого класса отмечается повышенная плотность костей по сравнению со спортсменами средней квалификации и, особенно, лицами, не занимающимися спортом. Представители скоростно-силовых видов спорта, вольной и греко-римской борьбы имеют достоверно более высокие показатели плотности костей по сравнению со спортсменами, специализирующимися в циклических, игровых и сложно-координационных видах спорта.
Большие объемы работы на выносливость приводят к снижению плотности костей (Michel et al., 1989). Особенно низкая плотность костей отмечается у пловцов на длинные дистанции, что обусловлено не только большим объемом работы аэробного характера, спецификой отбора пловцов, способных показать высокие результаты на стайерских дистанциях, но и спецификой водной среды, резко снижающей нагрузки на опорно-двигательный аппарат.

Не каждая нагрузка приводит к адаптационным перестройкам костной и волокнистой соединительной ткани. Чрезмерные нагрузки в отношении как их величины, так и продолжительности могут вызвать слишком большую функциональную деформацию, способную привести к травмам, которые возникают вследствие разовых перегрузок или в результате развившегося утомления (Carter, Carter, 1985). При незначительных нагрузках функциональная деформация недостаточна для развития адаптационных реакций костей, сухожилий и связок. Существенно также, что костная и волокнистые соединительные ткани проявляют нечувствительность к статическим нагрузкам, которые являются бесполезными для повышения их функциональных возможностей (Rubin, Lanyon, 1985; Zemicke, Loitz, 1992). Таким образом, рационально спланированные нагрузки являются действенным фактором укрепления и повышения функциональных возможностей костной, соединительной и хрящевой тканей. Под их влиянием увеличивается масса и плотность костей, улучшается транспорт питательных веществ и увеличивается плотность хряща, возрастает масса, эластичность и прочность (особенно в месте соединения с костью) связок, повышается эластичность и прочность сухожилий. В то же время чрезмерные механические силы, действующие на определенную структуру организма спортсмена, движения с чрезмерной амплитудой, высокая частота однонаправленных нагрузок могут не только нарушить процессы адаптации, но и привести к серьезным спортивным травмам (Нигг, 2002).
При наличии эффективных силовых раздражителей приспособительные изменения костной массы развиваются значительно медленнее, чем скелетной мускулатуры. Вместе с тем несколько лет напряженной силовой тренировки, характерной, например, для подготовки тяжелоатлетов, способны привести к очень значительным изменениям костной ткани (Stone, 1992). Установлено, что у юных квалифицированных тяжелоатлетов (средний возраст 17 лет) плотность минералов кости значительно превышает не только показатели контрольных испытуемых такого же возраста с незрелой костью, но и показатели взрослых мужчин со сформировавшейся костной массой (табл. 9.2).
Выявлена также достаточно тесная взаимосвязь между силовыми возможностями мышц бедер, спины и нижних конечностей производить силу и плотностью минералов проксимальной части бедренной кости и поясничного отдела позвоночного столба. Интересно отметить, что и чистая масса тела юных штангистов проявляет корреляционную связь с плотностью минералов кости. Это косвенно подтверждает тесную зависимость адаптационных перестроек кости от массы и силы мышц, поскольку от этих характеристик зависит воздействие мышечной массы на кость посредством сил, производимых сокращающейся мускулатурой. Установлено, что силовые нагрузки в детском и подростковом возрасте оказывают значительное влияние на плотность костей в зрелом возрасте (McCulloch et al., 1990). Молодая кость также очень восприимчива к внешним нагрузкам и реагирует на них интенсивным увеличением плотности минералов (Lanyon, 1987), что дает основания рекомендовать применение силовых упражнений с целью адаптации костной массы в пубертатном возрасте (Конрой и др., 1996).
Однако следует помнить, что в большинстве видов спорта большой объем напряженной силовой работы способен затормозить рост длинных костей и отрицательно повлиять на перспективы юных спортсменов (Matsuda et al., 1986), негативно воздействовать на механические характеристики костей, а также сухожилий и связок (Carter, 1984).
Говоря об адаптации костной ткани, следует отметить необходимость учета возрастных и половых особенностей спортсменов, а также особенностей питания, в частности потребления кальция.
В подростковом и юношеском возрасте, т. е. в период активного формирования скелета, рост костной массы во многом обусловливается рациональным питанием, содержащим в суточном рационе не только повышенное количество кальция (около 1200 м-сут~'), но и необходимое количество белков, фосфора, витаминов (особенно группы D). Это повышает эффективность кальциевого метаболизма и способствует увеличению костной массы и нормальному развитию скелета.
Интенсивно тренирующиеся спортсменки, особенно те из них, которые используют большие нагрузки в менструальный и постменструальный период, также нуждаются в кальциевой и гормональной поддержке с целью профилактики деминерализации костной ткани и развития остеопоро-за (Брукнер и др., 2002).
Адаптация сухожилий и связок к физическим нагрузкам включает большое количество различных изменений морфологического и биохимического характера. Длительная работа на развитие выносливости способствует интенсификации синтеза коллагена в сухожилиях (Stone, 1992). Силовая тренировка с большими отягощениями способствует увеличению содержания коллагена в связках и общего его содержания в соединительнотканных оболочках мышц, которые, являясь каркасом для передачи мышечной силы сухожилиям и костям, во многом определяют силу мышечного сокращения (Fleck, Falkel, 1986).
Важным итогом применения силовых упражнений является также повышение величины напряжения как в сухожилиях, так и в переходных элементах «кость — сухожилие», «кость — связка», «мышца — сухожилие» (Stone, 1992). Силовые упражнения, выполняемые с максимальной амплитудой и способствующие одновременному развитию силовых качеств и гибкости, являются эффективными для повышения длины и растяжимости сухожилий, накопления и увеличения силы за счет использования эластичных свойств сухожилий и соединительной ткани оболочек мышц.
Прекращение тренировки
и деадаптация мышечной ткани
Прекращение спортсменом интенсивной тренировки, особенно если оно связано с постельным режимом вследствие болезни или травмы, приводит к существенному снижению мышечной массы, которое отмечается уже через 3—4 дня. Более интенсивно процесс деадаптации затрагивает мышцы, которые подверглись интенсивным тренировочным воздействиям и являются наиболее значительными для того или иного вида спорта. Через 1,0—1,5 месяца постельного режима площадь поперечного сечения мышц может сократиться на 25-30 % (Fox et al., 1993).
Атрофии подвергаются все типы мышечных волокон. Однако наибольшее уменьшение мышечного объема отмечается в мышечных волокнах, которые были наиболее адаптированы в результате предшествовавшей тренировки. В процессе деадаптации может измениться и соотношение волокон различных типов. В частности, у спортсменов, специализирующихся в видах спорта, связанных с выносливостью к длительной работе, отличающихся высоким процентом мышечных МС-волокон и структурной и функциональной перестройкой БС-волокон, может отмечаться увеличение процента БСб- и БСа-волокон.
Процесс деадаптации не ограничивается мышечной тканью, он охватывает систему нервной регуляции движений. Проявляется это в том, что уменьшение мышечной силы вследствие неиспользования мышц возможно в большей мере, чем потеря объема мышечной массы. Это объясняется снижением способности нервной системы к рекрутированию мышечных единиц (Мак-Комас, 2001). Следствием атрофии мышц является и значительное увеличение их утомляемости, что связано со снижением возможностей всех звеньев систем энергоснабжения.
страница 1
|
Смотрите также:
В одной и той же мышце содержатся бс- и мс-волокна. Бс-волокна содержат активный фермент атфазу, который расщепляет атф с образованием больших количеств энергии, что обеспечивает быстрое сокращение волокон
346,06kb. 1 стр.
Материал, состоящий из мелких растительных волокон, соответствующим образом обработанных и соединенных в тонкий лист, в котором волокна связаны между собой поверхностными силами сцепления
32,64kb. 1 стр.
Утомление и восстановление в системе подготовки спортсменов
397,71kb. 1 стр.
|
|